sytidiini kuuluu nukleosideihin ja koostuu nukleiinipohjaisesta sytosiinista ja sokeririboosista. Se muodostaa emäsparin guanosiinin kanssa vety sidosten kautta. Sillä on myös keskeinen rooli pyrimidiiniaineenvaihdunnassa.
Mikä on sytidiini?
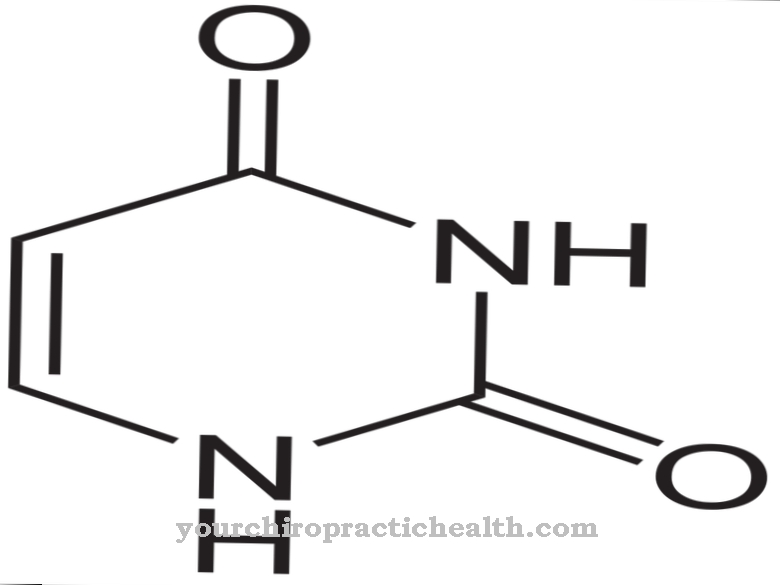
Sytidiini on nukleosidi, joka koostuu sytosiinista ja riboosista. Adeniinin, guaniinin ja tymiinin lisäksi typpipohjainen sytosiini osallistuu nukleiinihappojen synteesiin. Sytidiinin fosforylointi tuottaa sytidiinimonofosfaattia (CMP), sytidiinidifosfaattia (CDP) tai sytidiinitrifosfaattia (CTP).
Sytidiinimonofosfaatti on nukleotidi RNA: ssa. Kaksi puriini- ja kaksi pyrimidiiniemästä osallistuvat nukleiinihappojen rakenteeseen, ja tymiini vaihdetaan urasiiliksi RNA: ssa. Adeniini ja guaniini kuuluvat puriiniemäksiin, kun taas tymiini, sytosiini ja urasiili kuuluvat pyrimidiiniemäksiin. Sytidiinideaminaasi voi deaminoida sytidiinin uridiiniksi. Uridiini on nukleosidi, joka on valmistettu riboosista ja urasiilista. Se voidaan myös fosforyloida uridiinimonofosfaatiksi.
Uridiinimonofosfaatti on myös tärkeä nukleotidi RNA: lle. Lisäksi CDP ja CTP ovat myös aktivoivia ryhmiä lesitiinin, kefaliinin ja kardiolipiinin synteesille. Puhdas sytidiini on läsnä vesiliukoisena kiinteänä aineena, joka hajoaa lämpötilassa 201 - 220 astetta. Se voidaan hajottaa katalyyttisesti sytosiiniksi ja riboosiksi pyrimidiininukleosidaasientsyymin avulla.
Toiminto, vaikutukset ja tehtävät
Sytidiinillä on keskeinen rooli pyrimidiiniaineenvaihdunnassa. Pyrimidiini tarjoaa emäksisen rakenteen nukleiinihapoissa esiintyville sytosiini-, tymiini- ja urasiili-pyrimidiiniemäksille. RNA: ssa oleva tymiini vaihdetaan urasiiliin.
Urasiilia tuotetaan myös deaminoimalla sytidiini sytidiinideaminaasilla. Kemiallisilla muunnoksilla kolmen pyrimidiiniemäksen välillä on keskeinen merkitys DNA: n korjausprosesseissa ja epigeneettisissä muutoksissa. Epigenetian yhteydessä ympäristövaikutukset muuttavat erilaisia ominaisuuksia. Geneettinen materiaali ei kuitenkaan muutu. Organismin modifikaatiomuutokset johtuvat geenien erilaisesta ilmentymisestä. Kehossolujen erilaistumisprosessit eri solulinjojen ja elinten muodostamiseksi edustavat myös epigeneettistä prosessia. Solutyypistä riippuen eri geenit aktivoidaan tai deaktivoidaan.
Tämä tapahtuu sytidiiniemästen metyloinnin kautta DNA: ssa. Metyloinnin aikana muodostuu metyylisytosiini, joka voidaan muuttaa tymiiniksi deaminoimalla. Vastakkaisessa kaksoisjuosteessa oleva komplementaarinen nukleoabasaan guaniini mahdollistaa virheen tunnistamisen ja tymiinin vaihdon uudelleen sytosiiniksi. Kuitenkin guaniini voidaan myös vaihtaa adeniiniin, mikä johtaa pistemutaatioon. Jos metyloimaton sytosiini deaminoidaan, syntyy urasiilia. Koska urasiili ei esiinny DNA: ssa, se korvataan heti sytosiinilla. Sytosiinin sijasta metylaatiosta johtuva mutaationopeus kasvaa hieman.
Samanaikaisesti kuitenkin yhä useammat geenit kytketään pois käytöstä metyloinnin avulla, mikä johtaa solujen erikoistumiseen edelleen solulinjaan. Korjausprosesseissa korjausentsyymit perustuvat alkuperäiseen DNA-juosteeseen, jonka ne tunnistavat korkeammalla metylaatioasteella. Täydentävä osa on myös rakennettu siihen tallennettujen tietojen perusteella. Asennusvirheet korjataan välittömästi. Lisäksi entsyymi AID (aktivaation indusoima sytidiinideaminaasi) katalysoi spesifisesti sytidiiniryhmien deaminointia uridiiniryhmiksi yksijuosteisessa DNA: ssa. Somaattisia hypermutaatioita tapahtuu, jotka muuttavat B-solujen vasta-ainesekvenssejä. Sitten valitaan sopivat B-solut. Tämä mahdollistaa joustavan immuunivasteen.
Koulutus, esiintyminen, ominaisuudet ja optimaaliset arvot
Sytidiini on pyrimidiinimetabolian välituote. Eristettynä yhteytenä sillä ei ole merkitystä. Kuten jo mainittiin, se koostuu nukleiinipohjaisesta sytosiinista ja nelijalkaisesta sokeririboosista. Keho voi syntetisoida sytosiinin itse.
Sen synteesi on kuitenkin erittäin energiaintensiivinen, joten se otetaan talteen nukleiinihappojen rakennuspalikoista pelastuspolun puitteissa ja voidaan integroida uudelleen nukleiinihapoiksi. Kun emäs hajoaa kokonaan, syntyy hiilidioksidia, vettä ja ureaa. Se on läsnä nukleosidina RNA: ssa. DNA: ssa sytosiini on sitoutunut deoksiribosiiniin siten, että nukleosidideoksisitidiini esiintyy tässä rakennuspalikkana.
Sairaudet ja häiriöt
DNA: n sytidiinitähteiden metylaatiot ovat erittäin tärkeitä merkinnöille erilaisten biokemiallisten prosessien erottamiseksi. Methylaatiossa voi kuitenkin tapahtua virheitä, jotka johtavat sairauteen.
Virheellisissä metylaatioissa voidaan laukaista sekä lisääntynyt että vähentynyt geenitoiminta, joka ei täytä vaatimuksia. Nämä metylaatiomallit välittyvät solujen jakautumisen aikana. Pitkällä aikavälillä tapahtuu muutoksia, jotka voivat johtaa sairauksiin. Esimerkiksi joillakin tuumorisoluilla on erilaisia metylaatiorakenteita, joita ei esiinny terveissä soluissa. Esimerkiksi metylaatio voi estää tiettyjä geenejä, jotka koodaavat kasvua sääteleviä entsyymejä. Jos nämä entsyymit puuttuvat, solujen kasvu voi estyä. Tämä pätee myös entsyymeihin, jotka aloittavat solujen säännöllisen kuoleman (apoptoosi) soluvirheiden esiintyessä.
DNA-metyloinnin kohdennettu vaikuttaminen ei ole vielä mahdollista. Tuumorisolujen täydellisestä demetylaatiosta on kuitenkin tutkimuksia, jotta ne voitaisiin alistaa uudelleen kasvua säätelevien proteiinien hallintaan. Useiden kliinisten tutkimusten mukaan demetylaation on osoitettu rajoittavan tuumorin kasvua potilailla, joilla on akuutti myeloidileukemia. Tämä menetelmä tunnetaan myös nimellä epigeneettinen terapia. Metylaatioprosesseilla voi olla merkitys myös muissa sairauksissa. Ympäristövaikutusten takia organismi mukautuu muuttuneisiin olosuhteisiin muodostamalla biologisia muunnoksia, jotka perustuvat DNA: n sytidiinitähteiden metylaatioihin. Keho suorittaa siten oppimisprosessin, joka voi kuitenkin myös aiheuttaa väärän säätelyn.
.jpg)