aktiini on rakenteellinen proteiini, jota löytyy kaikista eukaryoottisoluista. Se osallistuu sytoskeleton ja lihaksen rakenteeseen.
Mikä on aktiini
Aktiini on evoluutiossa hyvin vanha proteiinimolekyyli. Rakenteellisena proteiinina se sisältyy jokaisen eukaryoottisolun sytoplasmaan ja kaikkien lihaskuitujen sarkomeeriin.
Yhdessä mikrotubulusten ja välifilamenttien kanssa se muodostaa jokaisen solun sytoskeletonin aktiinfilamenttien muodossa. Se vastaa yhdessä solurakenteen muodostumisesta ja molekyylien ja soluorganelien liikkeestä solun sisällä. Sama pätee kennojen yhteenkuuluvuuteen tiukkojen tai kiinnittyneiden liitosten kautta. Lihaskuiduissa aktiini yhdessä proteiinien myosiinin, troponiinin ja tropomyosiinin kanssa aiheuttaa lihasten supistumisia.
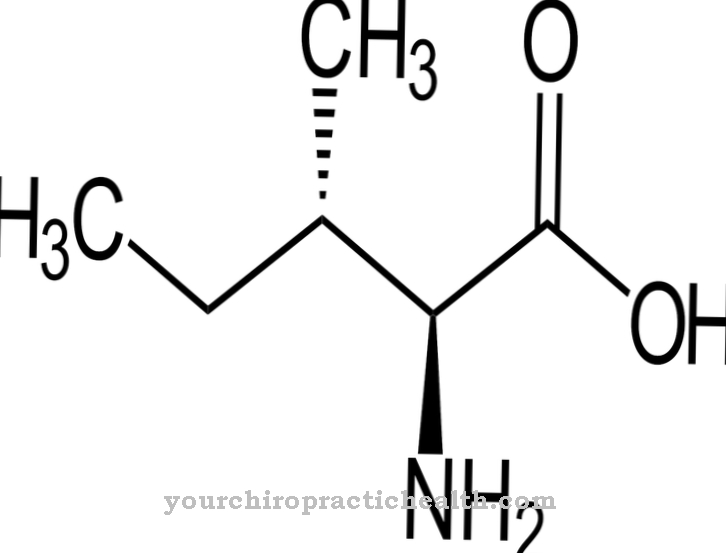
Aktiini voidaan jakaa kolmeen funktionaaliseen yksikköön: alfa-aktiini, beeta-aktiini ja gammaktiini. Alfa-aktiini on lihaskuitujen rakenteellinen komponentti, kun taas beeta- ja gammaktiinia löytyy pääasiassa solujen sytoplasmasta. Aktiini on erittäin konservoitunut proteiini, jota esiintyy yksisoluisissa eukaryoottisoluissa erittäin pienillä poikkeamilla aminohapposekvenssissä. Ihmisillä 10 prosenttia kaikista lihassolujen proteiinimolekyyleistä koostuu aktiinista. Kaikki muut solut sisältävät edelleen 1 - 5 prosenttia tästä molekyylistä sytoplasmassa.
Toiminto, vaikutukset ja tehtävät
Aktiini täyttää tärkeät toiminnot soluissa ja lihaskuiduissa. Solun sytoplasmassa osana sytoskelettoa se muodostaa tiheän, kolmiulotteisen verkon, joka pitää solurakenteet yhdessä.
Tietyissä verkon kohdissa rakenteet vahvistuvat muodostaen kalvon pullistumia, kuten mikrovilli, synapses tai pseudopodia. Adherens-liitokset ja tiukka liitososat ovat saatavissa solukontaktille. Kaiken kaikkiaan aktiini edistää solujen ja kudosten vakautta ja muotoa. Stabiinin lisäksi aktiini varmistaa myös kuljetusprosessit solun sisällä. Se sitoo tärkeitä rakenteellisesti sukulaisia kalvon läpäiseviä proteiineja siten, että ne pysyvät lähellä. Mioosiinien (motoriset proteiinit) avulla aktiinikuidut myös kuljettavat lyhyitä matkoja.
Esimerkiksi vesikkelit voidaan kuljettaa kalvoon. Pikemmät jaksot otetaan mikrotubulusten toimesta motoristen proteiinien kinesiinin ja dyneiinin avulla. Aktiini varmistaa myös solujen liikkuvuuden. Solujen on voitava liikkua kehossa monissa tapauksissa. Tämä koskee erityisesti immuunireaktioita tai haavan paranemista, samoin kuin solujen muodon yleisiä liikkeitä tai muutoksia. Liikkeet voivat perustua kahteen eri prosessiin. Yhtäältä liike voidaan laukaista suunnatulla polymerointireaktiolla ja toisaalta aktiini-myosiini-vuorovaikutuksen kautta.
Aktiini-myosiini-vuorovaikutuksessa aktiinikuidut muodostuvat fibrillipaketeiksi, jotka toimivat kuten köyttä vetämällä myosiinin avulla. Aktiinilangat voivat aiheuttaa solujen kasvua pseudopodian (filopodia ja lamellipodia) muodossa. Solussa olevien monimuotoisten toimintojen lisäksi aktiini on tietysti vastuussa sekä luurankolihasten että sileiden lihasten lihaksen supistumisesta. Nämä liikkeet perustuvat myös aktiini-myosiini-vuorovaikutukseen. Tämän varmistamiseksi monet aktiinifilamentit on kytketty muihin proteiineihin erittäin järjestetyllä tavalla.
Koulutus, esiintyminen, ominaisuudet ja optimaaliset arvot
Kuten jo mainittiin, aktiinia löytyy kaikista eukaryoottisista organismeista ja soluista. Se on sytoplasman luontainen osa ja varmistaa solujen stabiilisuuden, rakenteellisesti samankaltaisten proteiinien ankkuroinnin, vesikkelien lyhyen matkan kuljetuksen solukalvoon ja solun liikkuvuuden. Ilman aktiinia solu ei voisi selviytyä. Aktiinivariantteja on kuusi, jotka on jaettu kolmeen alfavarianttiin, yhteen beetavarianttiin ja kahteen gammavarianttiin.
Alfa-aktiinit osallistuvat lihaksen kehitykseen ja supistumiseen. Beeta-aktiinilla ja gamma-1-aktiinilla on suuri merkitys sytoplasman sytoskeletonille. Gamma-2-aktiini puolestaan vastaa sileistä lihaksista ja suoliston lihaksista. Synteesin aikana muodostuu aluksi monomeerinen globaali aktiini, joka tunnetaan myös nimellä G-aktiini. Yksittäiset monomeeriset proteiinimolekyylit puolestaan polymeroituvat filamentaarisen F-aktiinin muodostamiseksi.
Polymerointiprosessin aikana useat pallomaiset monomeerit yhdistyvät muodostaen pitkän, säikemäisen F-aktiinin. Sekä ketjujen rakenne että hajoaminen ovat erittäin dynaamisia. Tällä tavoin aktiinikehys voidaan mukauttaa nopeasti nykyisiin vaatimuksiin. Lisäksi tämä prosessi varmistaa myös solujen liikkeet. Näitä reaktioita voidaan estää ns. Sytoskeletaaliset estäjät. Näillä aineilla joko polymeroituminen tai depolymeroituminen estyy. Ne ovat lääketieteellisesti tärkeitä lääkkeinä kemoterapian yhteydessä.
Sairaudet ja häiriöt
Koska aktiini on olennainen osa kaikkia soluja, monet mutaatioiden aiheuttamat rakennemuutokset johtavat organismin kuolemaan. Alfa-aktiinien geenien mutaatiot voivat aiheuttaa lihassairauksia. Tämä pätee erityisesti alfa-1-aktiiniin.
Koska alfa-2-aktiini on vastuussa aortan lihaksista, perinnöllinen rintakehän aortan aneurysma voi ilmetä, jos ACTA2-geeni on mutatoitunut. ACTA2-geeni koodaa alfa-2-aktiinia. Sydämen alfaaktiinin mutaatio ACTC1-geenissä aiheuttaa laajentuneen kardiomyopatian. Lisäksi ACTB: n mutaatio sytoplasmisen beeta-aktiinin geeninä voi aiheuttaa suurten solujen ja diffuusion B-solujen lymfooman. Joillakin autoimmuunisairauksilla voi olla kohonneet aktiinivasta-aineiden tasot.
Tämä koskee erityisesti autoimmuunista maksatulehdusta. Se on krooninen hepatiitin kulku, joka johtaa maksakirroosiin pitkällä aikavälillä. Täältä löytyy vasta-aine sileän lihaksen aktiinia vastaan. Erodiagnoosin kannalta autoimmuuninen hepatiitti ei ole niin helppo erottaa kroonisesta virushepatiitista. Koska kroonisessa virushepatiitissa myös aktiinivasta-aineita voidaan stimuloida vähemmässä määrin.
.jpg)

.jpg)
.jpg)







.jpg)


.jpg)