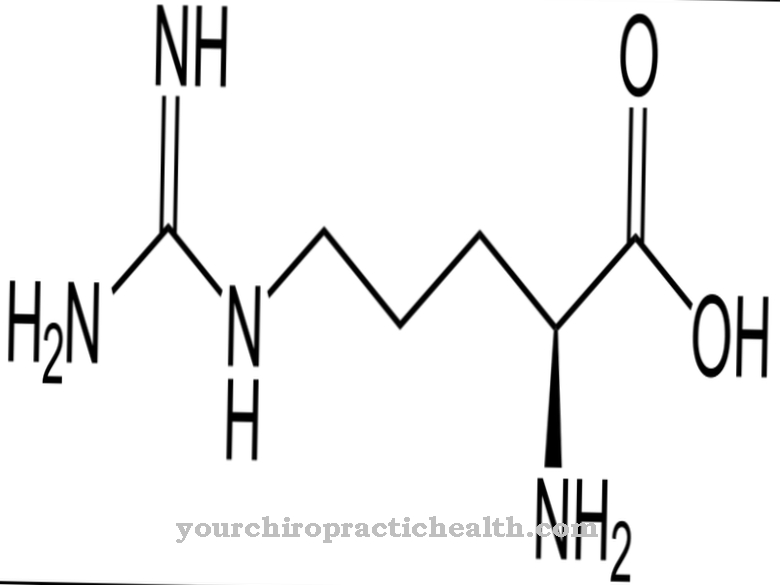
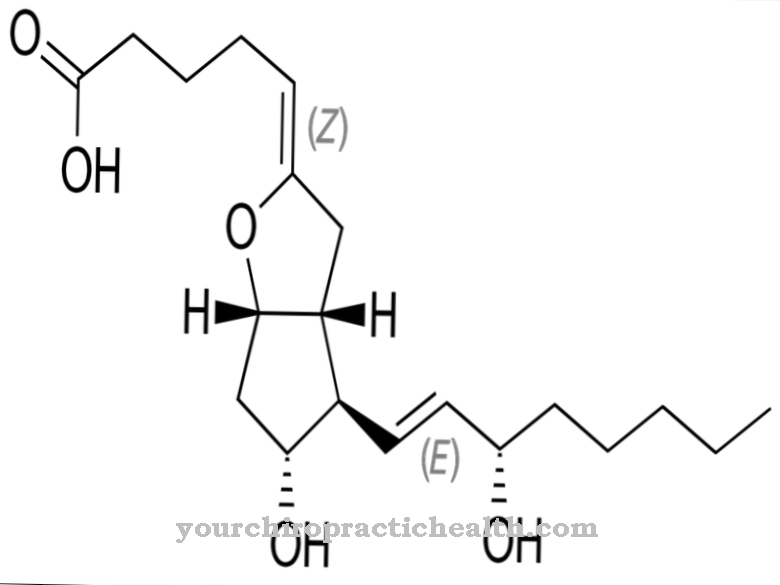
nukleoemäksestä ovat rakennuspalikoita, joista DNA: n ja RNA: n pitkät ketjut koostuvat fosforyloidussa nukleotidimuodossaan.
DNA: ssa, joka muodostaa kaksoisjuosteet, jotka ovat samanlaisia kuin köydet, 4 esiintyvää nukleiiniemästä muodostavat kiinteitä pareja vastaavan komplementaarisen emäksen kanssa vetysiltojen kautta. Nukleobaasit koostuvat joko bisyklisestä puriinista tai monosyklisestä pyrimidiinirungosta.
Mitä ovat nukleoalukset?
4 nukleaabasaasia adeniini, guaniini, sytosiini ja tymiini muodostavat pitkien DNA-kaksoisheeliksimolekyyliketjujen rakennuspalikoina vakioparit adeniini-tymiini (A-T) ja guaniini-sytosiini (G-C).
Kaksi emästä adeniini ja guaniini koostuvat kumpikin modifioidusta bisyklisestä kuuden ja viiden jäsenen renkaasta, jolla on emäksinen puriinirakenne, ja siksi niitä kutsutaan myös puriiniemäksiksi. Kahden muun nukleiiniemäksen, sytosiinin ja tymiinin, perusrakenne koostuu heterosyklisestä aromataisesta kuusijäsenisestä renkaasta, joka vastaa modifioitua pyrimidiinirunkoa, minkä vuoksi niihin viitataan myös pyrimidiiniemäksinä. Koska RNA on pääosin läsnä yksittäisissä juosteissa, emäsparia ei aluksi ole. Tämä tapahtuu vain replikaation aikana mRNA: n (messenger RNA) kautta.
RNA-juosteen kopio koostuu komplementaarisista nuklea-emäksistä, jotka ovat analogisia DNA: n toisen juosteen kanssa. Ainoa ero on, että urasiili korvataan tymiinillä RNA: ssa. DNA- ja RNA-ketjun molekyylit eivät muodostu nukleobaasien puhtaassa muodossa, vaan DNA: n tapauksessa ne yhdistyvät 5-sokerin deoksiribosiinin kanssa vastaavan nukleosidin muodostamiseksi. RNA: n tapauksessa sokeriryhmä koostuu riboosista. Lisäksi nukleosidit fosforyloidaan fosfaattijäännöllä niin kutsuttujen nukleotidien muodostamiseksi.
Puriiniemäkset hypoksantiini ja ksantiini, joita esiintyy myös DNA: ssa ja RNA: ssa, vastaavat modifioitua tymiiniä. Hypoksantiini muodostuu adeniinista korvaamalla aminoryhmä (-NH3) hydroksyyliryhmällä (-OH), ja ksantiinia muodostuu guaniinista. Molemmat nuklea-emäkset eivät edistä geneettisen tiedon välitystä.
Toiminto, vaikutukset ja tehtävät
Yksi tärkeimmistä DNA: n kaksoisjuosia muodostavien nukleobaasien toiminnoista on osoittaa läsnäolo halutussa paikassa.
Nukleobaasien sekvenssi vastaa geneettistä koodia ja määrittelee aminohappojen tyypin ja sekvenssin, josta proteiinit koostuvat. Tämä tarkoittaa, että nukleobaasien tärkein tehtävä osana DNA: ta on passiivinen, staattinen rooli, ts. Ne eivät puutu aktiivisesti aineenvaihduntaan ja lähetti-RNA (mRNA) ei muuta niiden biokemiallista rakennetta lukuprosessin aikana. Tämä selittää osittain DNA: n pitkäikäisyyden.
Mitokondriaalisen DNA: n (mtDNA) puoliintumisaika, jonka aikana puoli alun perin olemassa olevista sidoksista hajoaa, on suuresti riippuvainen ympäristöolosuhteista ja vaihtelee noin 520 vuoden välillä keskimääräisissä olosuhteissa positiivisissa lämpötiloissa ja jopa 150 000 vuoteen ikirouta-olosuhteissa. .
Osana RNA: ta nuklea-emäkset pelaavat jonkin verran aktiivisempaa roolia. Periaatteessa solujen jakautuessa DNA-kaksoisjuosteet hajotetaan ja erotetaan toisistaan komplementaarisen juosteen, mRNA: n, muodostamiseksi, joka niin sanotusti muodostaa geneettisen materiaalin työkopion ja toimii perustana niiden aminohappojen valinnalle ja sekvenssille, joista aiotut proteiinit kootaan. Toinen nukleiiniemäs, dihydrourasiili, esiintyy vain ns. Kuljetus-RNA: ssa (tRNA), jota käytetään aminohappojen kuljettamiseen proteiinisynteesin aikana.
Jotkut nuklea-emäkset suorittavat täysin erilaisen funktion osana entsyymejä, jotka mahdollistavat ja katalysoivat aktiivisesti tiettyjä biokemiallisia prosesseja. Adeniini suorittaa tunnetuimman tehtävänsä nukleotidina solujen energiatasapainossa. Adeniinilla on tärkeä rooli elektronien luovuttajana adenosiinidifosfaattina (ADP) ja adenosiinitrifosfaattina (ATP), samoin kuin nikotiiniamidi-adeniinidinukleotidin (NAD) komponenttina.
Koulutus, esiintyminen, ominaisuudet ja optimaaliset arvot
Fosforyloimattomassa muodossa nuklea-emäkset koostuvat yksinomaan hiilestä, vedystä ja hapesta, aineista, jotka ovat kaikkialla läsnä olevia ja vapaasti saatavissa olevia. Keho pystyy siis syntetisoimaan nukleobaaseja itse, mutta prosessi on monimutkainen ja energiaintensiivinen.
Siksi nukleiinihappojen talteenotto kierrätyksellä on edullista, esim. B. hajottamalla proteiineja, jotka sisältävät tiettyjä yhdisteitä, jotka voidaan eristää ja muuntaa nukleiinihapoiksi vähäisin energiankulutuksin tai jopa energian voitolla. Nukleiinihappoja ei yleensä esiinny puhtaassa muodossa kehossa, vaan enimmäkseen nukleosideina tai deoksinukleosideina, joihin on kiinnittynyt riboosi- tai deoksiribosimolekyyli. DNA: n ja RNA: n komponenttina ja tiettyjen entsyymien komponenttina nukleiinihapot tai niiden nukleosidit fosforyloituvat myös palautuvasti yhdestä kolmeen fosfaattiryhmällä (PO4-).
Nukleobaasien optimaaliselle tarjonnalle ei ole viitearvoa. Nukleobaasien puute tai ylimäärä voidaan määrittää vain epäsuorasti tiettyjen aineenvaihdunnan häiriöiden kautta.
Sairaudet ja häiriöt
Sellaiset vaarat, häiriöt ja riskit, jotka tuodaan yhteyteen nukleobaasien kanssa, ovat DNA- tai RNA-juosteiden lukumäärän ja sekvenssin virheitä, jotka johtavat muutokseen proteiinisynteesin koodauksessa.
Jos elimistö ei pysty korjaamaan virhettä korjausmekanismiensa avulla, syntyy biologisesti inaktiivisten tai käyttökelpoisten proteiinien synteesi, mikä puolestaan voi johtaa lievistä tai vakaviin aineenvaihduntahäiriöihin. Se voi esimerkiksi B. geenimutaatioita on läsnä, jotka voivat laukaista oireellisia sairauksia alusta alkaen metabolisten häiriöiden kautta, jotka voivat olla parantumattomia. Mutta jopa terveessä genomissa, kopiointivirheitä voi tapahtua DNA- ja RNA-ketjujen replikaation aikana, jotka vaikuttavat aineenvaihduntaan.
Tunnettu metabolinen häiriö puriinitasapainossa on z. B. takaisin geneettiseen vikaan x-kromosomissa. Geneettisen vian takia puriiniemäksiä, hypoksantiinia ja guaniinia ei voida kierrättää, mikä edistää viime kädessä virtsakivien ja kihdin muodostumista niveliin.
.jpg)


.jpg)
.jpg)







.jpg)


.jpg)